Acoplamiento excitación-contracción
El acoplamiento excitación-contracción, o ECC, es el proceso mediante el cual una señal eléctrica en una célula muscular hace que esa célula muscular se acorte mecánicamente. Como sugiere su nombre, la ECC consta de dos fases vinculadas: la fase de excitación y la fase de contracción.
La excitación se refiere a la generación de un potencial de acción dentro de la célula muscular. Un potencial de acción es un evento eléctrico, lo que significa que implica un cambio de carga. La señal eléctrica que conduce a la contracción de las células musculares comienza como un impulso nervioso y se transmite a los músculos a través de neuronas motoras. La conexión entre una neurona motora y una célula muscular se llama unión neuromuscular. Aquí, los impulsos se transmiten entre el sistema nervioso y los músculos, propagando el potencial de acción que conducirá a la contracción muscular.
La contracción se refiere a un cambio físico en la forma de la célula muscular (también llamada fibra muscular ). Cuando una fibra muscular se contrae, se acorta. Los músculos están formados por muchas fibras musculares altamente organizadas; cuando se contraen al unísono, la forma de todo el músculo cambia para volverse más corto y flexionado.
Diagrama de acoplamiento excitación-contracción en el músculo esquelético y cardíaco
Los músculos están formados por haces de fibras musculares altamente organizados. Dentro de un músculo, grupos de fibras musculares se agrupan en asociaciones llamadas fascículos. La membrana encargada de mantener unidos los fascículos se llama perimisio. El músculo en sí consta de múltiples fascículos agrupados, con vasos sanguíneos que suministran oxígeno y neuronas motoras para enviar impulsos eléctricos. Una vaina llamada epimisio es la capa exterior que mantiene unidos estos componentes.
Incluso a nivel celular, las células musculares están muy organizadas. Una célula muscular es una fibra larga y delgada. Cada uno contiene un orgánulo que es exclusivo de las células musculares, llamado retículo sarcoplásmico o SR. La función principal de este orgánulo es almacenar iones de calcio, que son vitales para la CEC. El SR está estrechamente asociado con pequeñas fibras, llamadas miofibrillas, que constituyen la mayor parte de la célula muscular. Las miofibrillas consisten en regiones superpuestas de miosina o filamentos gruesos y actina o filamentos delgados.
Glándulas Mamarias: Anatomía, función y diagrama
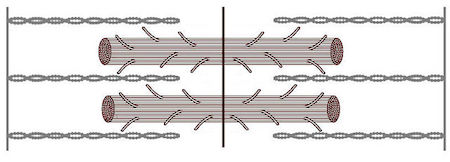 |
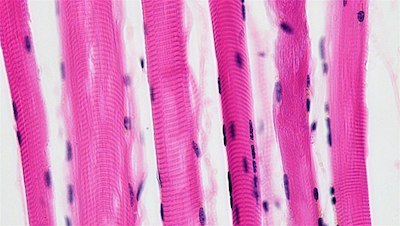
En los músculos esqueléticos, la miosina y la actina están organizadas de forma intrincada. Cada conjunto de una banda gruesa de miosina y una banda delgada de actina se llama sarcómero. El patrón repetitivo de disposición de los sarcómeros dentro del músculo esquelético da a estas células musculares una apariencia rayada o estriada; por esta razón, los músculos esqueléticos se denominan «músculos estriados».
 |
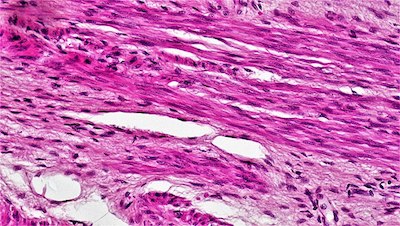
Esto es diferente del tipo de células musculares que se encuentran en las paredes del sistema digestivo, en las que hay menos organización de actina y miosina. Sin sarcómeros, estas células carecen de la apariencia rayada de los músculos esqueléticos y, por lo tanto, se las denomina «músculo liso».
 |
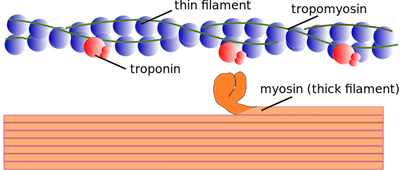
La actina y la miosina se conocen como proteínas contráctiles, lo que significa que canalizan la energía generada a través de la hidrólisis del ATP para provocar la contracción cinética real de las células musculares. En el sistema actina/miosina, la miosina actúa como una proteína motora y la actina como una pista estructural que sigue el motor de miosina. A través de la unión repetida del motor de miosina a sucesivos sitios activos de la vía de actina, este dúo de proteínas provoca la contracción muscular.
Si bien son importantes, las proteínas contráctiles no son los únicos participantes proteicos en la CEC. Para este proceso también son esenciales las proteínas reguladoras, que controlan y dirigen la acción de las proteínas contráctiles. La tropomiosina y la troponina son dos ejemplos importantes de proteínas reguladoras. Estas proteínas se unen a los sitios activos de los filamentos de actina, evitando que la miosina se una y avance la contracción muscular. Por tanto, la presencia de tropomiosina y troponina en los filamentos de actina puede impedir el movimiento muscular.
 |
Considere tres pasos del acoplamiento excitación-contracción, comenzando con la excitación eléctrica.
¿Cuál es el propósito de la meditación en el budismo?
El movimiento muscular comienza en el sistema nervioso cuando un impulso eléctrico se propaga y se envía a través de una neurona motora a la unión neuromuscular, el sitio de comunicación química entre una neurona y una célula muscular. En la unión, la neurona excitada libera el neurotransmisor acetilcolina en la hendidura sináptica, donde se une a receptores en la superficie de la célula muscular e inicia un potencial de acción, un rápido cambio de polarización a través de la membrana.
El potencial de acción viaja rápidamente a lo largo de la fibra muscular. Para permitir que el potencial de acción alcance incluso las miofibrillas que están enterradas más profundamente dentro de la célula, la membrana celular de una fibra muscular tiene muchas invaginaciones pequeñas en forma de tubo que llegan hasta el citosol. Estos se denominan «túbulos transversales» o túbulos en T. Los túbulos T funcionan para transportar potenciales de acción más profundamente al interior de la célula.
A medida que el potencial de acción viaja a lo largo del túbulo T, los receptores detectores de voltaje en la membrana registran el cambio resultante en el voltaje de la membrana. En respuesta, estos receptores (llamados «receptores de dihidropiridina») sufren un cambio conformacional e interactúan alostéricamente con otra proteína muy próxima: los receptores de rianodina en la membrana del retículo sarcoplásmico. Esta interacción abre los canales iónicos de calcio en el SR, lo que provoca que la célula se inunde con Ca2+.
Estos iones de calcio son necesarios para la contracción muscular en la segunda fase de la ECC. El Ca2+ que inunda el citosol de las células musculares tiene una alta afinidad por la troponina. Los iones están presentes en una concentración tan alta y tienen una afinidad tan alta por la troponina que se unen inmediatamente, provocando un cambio conformacional que desmonta el complejo troponina-actina.
Con las proteínas reguladoras ahora separadas de la actina, pasamos a la tercera fase de la ECC. El complejo motor actina-miosina ahora puede formarse libremente y continuará la contracción mecánica. Cuando la ECC se produce en todo un músculo, las contracciones de tantas células musculares hacen que el músculo se contraiga y se flexione.
Ciclismo de puentes cruzados y calcio en la contracción muscular
La capacidad contráctil del complejo miosina-actina se debe a un proceso repetitivo llamado ciclo de puentes cruzados. Al comienzo de este proceso, las cabezas activas de miosina se unen al ATP. Cuando esta molécula de ATP se hidroliza para convertirse en ADP, la molécula de miosina puede unirse con la actina adyacente. Luego, la molécula de ADP es expulsada de la miosina. Esta reacción provoca el golpe de poder de la miosina, un movimiento de trinquete mediante el cual la miosina arrastra las moléculas de actina más cerca del centro del sarcómero.
Ahora, una nueva molécula de ATP encuentra y se une a la molécula de miosina. La unión del ATP hace que la miosina se desprenda de la actina y se completa una ronda del ciclo de puentes cruzados. Para que se produzca el ciclo de puentes cruzados, el ATP debe estar presente en la célula en grandes cantidades para que la miosina pueda unirse y separarse repetidamente de la actina, acortando continuamente el sarcómero.
En el músculo liso, una proteína llamada calmodulina ayuda a activar la contracción. En respuesta a la fluctuación de los iones calcio, la calmodulina activa la unión del ATP a la miosina, lo que provoca la contracción muscular.
El modelo de filamento deslizante de excitación-contracción en el músculo esquelético
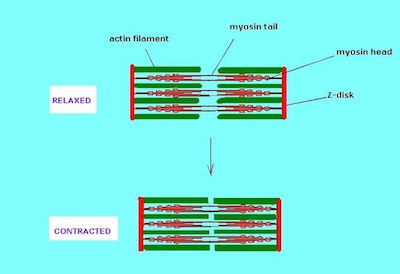 |
Este diagrama representa el modelo de filamento deslizante de contracción muscular en una célula de músculo esquelético. En este modelo, la miosina (roja) y la actina (verde) se colocan una encima de la otra en un patrón alterno. En reposo, los filamentos de miosina y actina pueden deslizarse entre sí, alargando la fibra muscular.
Sin embargo, durante el acoplamiento de excitación-contracción en el músculo esquelético, la interacción entre actina y miosina hace que los filamentos de actina se junten a lo largo de las fibrillas de miosina. Este movimiento de deslizamiento hace que el sarcómero se acorte y, en última instancia, el músculo se contraiga.
El proceso de contracción y relajación muscular
¿Qué estimula la relajación de los músculos? De hecho, es la ausencia de una señal, más que la estimulación directa, lo que hace que los músculos dejen de contraerse. Cuando el sistema nervioso deja de enviar impulsos para contraerse, no se propagan potenciales de acción en la fibra muscular. Esto permite que la membrana celular, incluidos los túbulos T, se repolarice. Los canales de calcio dependientes del voltaje lo detectan y se cierran.
Una bomba de iones de calcio en el SR (llamada «ATPasa cálcica del retículo sarcoendoplasmático» o SERCA) elimina rápidamente Ca2+ del espacio citosólico, secuestrándolo una vez más dentro del SR. El ATP disponible se unirá a la miosina, provocando que se separe del ATP. Pero en ausencia de calcio, las proteínas reguladoras como la troponina y la tropomiosina se unen una vez más a la actina, protegiéndola de la unión de la miosina hasta que otro potencial de acción comienza de nuevo el ciclo.
Después de la muerte, el cuerpo deja de producir ATP. Un efecto de esto es la obstrucción de este ciclo: sin ATP, la miosina no puede separarse de la actina. Esto hace que los músculos se flexionen permanentemente en la condición rígida y rígida conocida como rigor mortis, que puede comenzar tan pronto como dos a cuatro horas después de que se haya producido la muerte.
Fuentes de ATP
¿De dónde viene el ATP tan importante para la ECC? Existen varias fuentes de ATP utilizadas por el tejido muscular en diferentes fases de contracción.
El fosfato de creatina es una molécula de alta energía que puede generar más ATP transfiriendo su grupo fosfato a una molécula de ATP. El fosfato de creatina está presente en grandes cantidades en el tejido muscular y ayuda a potenciar la contracción muscular durante aproximadamente los primeros quince segundos.
Una vez que se agota el fosfato de creatina, los músculos pueden utilizar la glucólisis para generar ATP. En este proceso, la descomposición de una molécula de glucosa se puede realizar incluso en ausencia de oxígeno para producir ATP y ácido pirúvico. Sin embargo, produce ATP más lentamente que el uso de fosfato de creatina.
El ácido pirúvico de la glucólisis también puede producir ATP. Si el sistema circulatorio suministra suficiente oxígeno al tejido muscular, el piruvato se descompondrá aeróbicamente en las mitocondrias para producir ATP adicional y el producto de desecho del dióxido de carbono. Sin embargo, si no hay suficiente oxígeno (por ejemplo, en caso de esfuerzo extremo), el piruvato también puede descomponerse anaeróbicamente. Este proceso, llamado metabolismo del ácido láctico, hace que el ácido láctico se acumule en el tejido muscular y puede provocar fatiga muscular.
Resumen de la lección
El acoplamiento excitación-contracción, o ECC, es el proceso que vincula las señales eléctricas con el movimiento mecánico del cuerpo. La ECC consta de una fase de excitación y una fase de contracción. Durante la excitación, se propaga un potencial de acción en la membrana de la célula muscular. Este potencial de acción desencadena la liberación de iones de calcio dentro de la célula, iniciando la contracción.
Dentro de las células musculares individuales hay numerosas fibras largas, llamadas miofibrillas, compuestas por dos proteínas: miosina y actina. Estas proteínas se conocen como proteínas contráctiles por su implicación en la contracción mecánica de la célula muscular. Un orgánulo llamado retículo sarcoplásmico también está presente dentro de las células musculares y es responsable de secuestrar iones de calcio cuando no hay potencial de acción presente.
Durante la ECC, el potencial de acción hace que la célula muscular se inunde de calcio. Esto libera proteínas reguladoras como la troponina y la tropomiosina de sus sitios de unión inhibidoras en la actina. Luego, a través de la unión e hidrólisis repetidas del ATP sobre la miosina, los filamentos de miosina y actina se mueven uno sobre otro en el modelo de contracción muscular de filamento deslizante. Esto acorta la fibra muscular y se llama ciclo de puente cruzado. Cuando se detiene la estimulación muscular del sistema nervioso, el SR vuelve a secuestrar iones de calcio. El ATP se une a la miosina, lo que hace que libere actina y el músculo se relajará hasta que se estimule una vez más.
Explora más sobre este tema
Selecciona un tema y sigue aprendiendo...
