Cuando pensamos en el mecanismo que sostiene la vida en la Tierra, la mente nos transporta de forma casi automática a un escenario cubierto de vegetación. Imaginamos bosques tropicales densos, campos de cultivo que se extienden hasta el horizonte o el sutil vaivén del fitoplancton bajo la superficie del océano. En todos estos paisajes se desarrolla el mismo prodigio biológico: las plantas absorben luz solar, consumen dióxido de carbono, toman agua del sustrato y, mediante un despliegue de microingeniería evolutiva, liberan bocanadas de oxígeno puro a la atmósfera. Este fenómeno ha dominado la narrativa científica escolar durante generaciones. Sin embargo, en los rincones más profundos, oscuros y hostiles de nuestro planeta, donde el oxígeno es un veneno mortífero y la luz solar apenas llega como un susurro, opera una versión alternativa, arcaica y fascinante de este proceso: la fotosíntesis anoxigénica.
Este canal metabólico representa un viaje al pasado geológico de la Tierra, un testimonio vivo de cómo funcionaba la biosfera miles de millones de años antes de que el primer brote verde emergiera del suelo. Lejos de ser una mera curiosidad de laboratorio, este sistema de nutrición molecular es ejecutado por comunidades de bacterias especializadas que habitan en lodos sulfurosos, lagos salinos de gran profundidad y fuentes hidrotermales submarinas. Al adentrarnos en su funcionamiento, descubrimos que la luz puede transformarse en energía química sin necesidad de producir una sola molécula de oxígeno, utilizando en su lugar compuestos tan singulares como el azufre o el hidrógeno gaseoso. Explorar este universo microscópico nos obliga a redefinir los límites de la vida y nos ofrece pistas cruciales sobre cómo interactúan la geología y la biología en mundos distantes.
La bifurcación de los senderos fotótrofos
Para comprender la esencia de esta modalidad metabólica, es indispensable desarmar el concepto general de fototrofia. La capacidad de capturar la radiación electromagnética del Sol para edificar materia orgánica no sigue un único camino evolutivo. La naturaleza bifurcó este proceso en dos grandes estrategias que conviven en el planeta, diferenciadas no por la fuente de energía que emplean, sino por las materias primas que eligen para saldar sus deudas electrónicas.
El modelo tradicional oxigénico
La versión más difundida y conocida de la fotosíntesis utiliza el agua ({eq}\text{H}_2\text{O}{/eq}) como el donador fundamental de electrones. Las plantas y las cianobacterias absorben la luz solar para romper la molécula de agua, un proceso de fotólisis que arranca los electrones necesarios para reducir el dióxido de carbono y fabricar carbohidratos. Como el agua está compuesta por hidrógeno y oxígeno, al despojarla de sus electrones y protones, el átomo de oxígeno remanente queda libre, se asocia con otro átomo idéntico y se expulsa a la atmósfera en forma de gas ({eq}\text{O}_2{/eq}). Esta estrategia es la responsable directa de que nuestra atmósfera sea respirable en la actualidad.
La alternativa desprovista de oxígeno

La fotosíntesis anoxigénica, por el contrario, prescinde por completo del agua como fuente de electrones. Los organismos que la ejecutan habitan en entornos donde el agua, aunque abundante, no es explotada de ese modo debido a las limitaciones de sus maquinarias moleculares. En su lugar, estas bacterias recurren a compuestos inorgánicos reducidos que se encuentran en su entorno inmediato, como el sulfuro de hidrógeno ({eq}\text{H}_2\text{S}{/eq}), el hidrógeno molecular ({eq}\text{H}_2{/eq}) o incluso el hierro ferroso ({eq}\text{Fe}^{2+}{/eq}).
Al oxidar estas sustancias para extraer sus electrones, el subproducto resultante jamás será oxígeno gaseoso. Si el donador inicial fue sulfuro de hidrógeno, el residuo acumulado en el medio será azufre elemental sólido, el cual se deposita en forma de diminutos gránulos brillantes dentro o fuera de la célula bacteriana.
[Fotosíntesis Oxigénica]: Luz + CO2 + H2O -------> Glucosa + O2 (Oxígeno) [Fotosíntesis Anoxigénica]: Luz + CO2 + H2S -------> Glucosa + S (Azufre Sólido) La maquinaria interna: Pigmentos del inframundo espectral
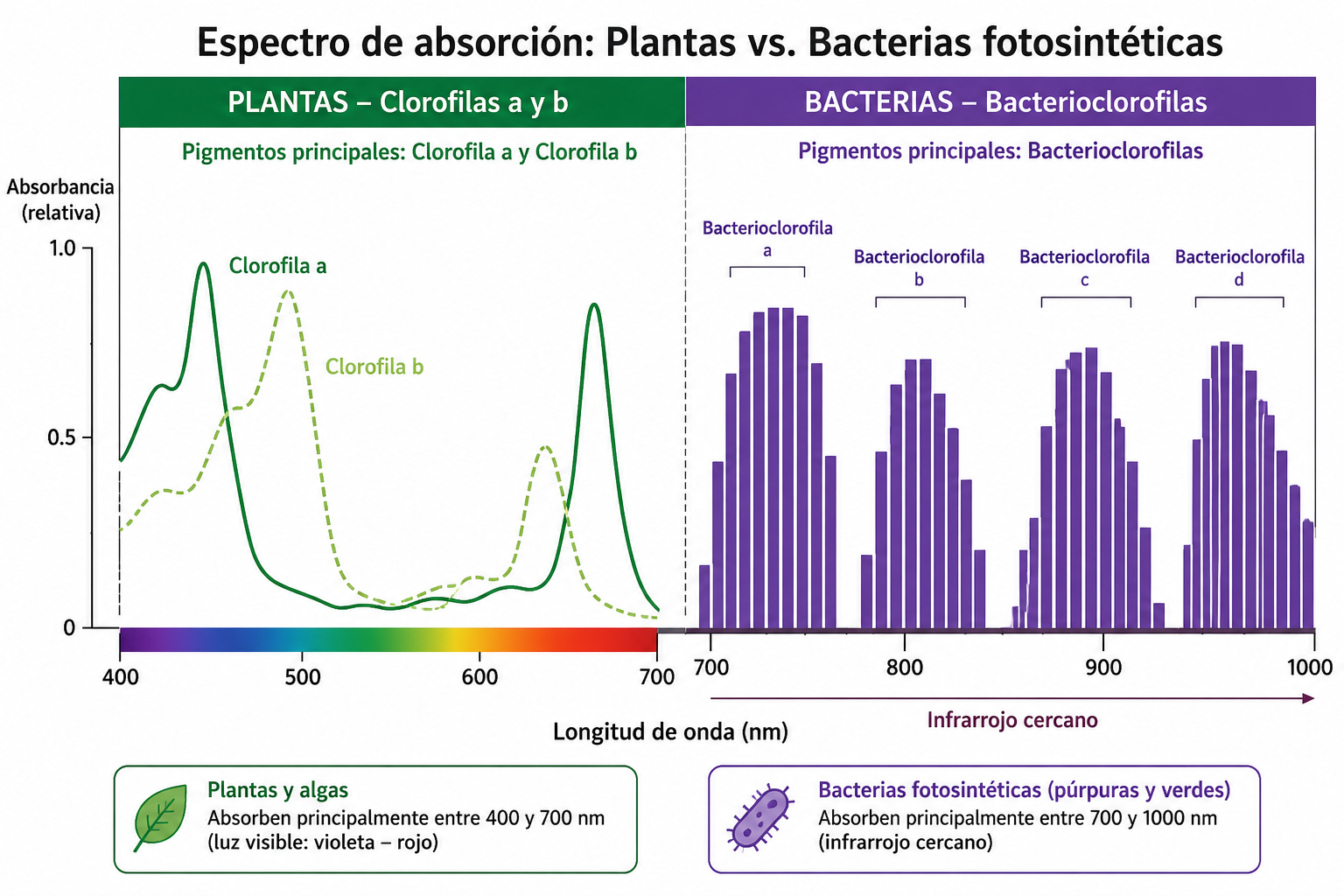
La captación de energía lumínica requiere antenas moleculares capaces de sintonizar las frecuencias específicas de la radiación solar. Mientras que los organismos verdes del mundo exterior confían sus tareas a las clorofilas convencionales, las bacterias anoxigénicas han desarrollado una familia de pigmentos única, diseñada para operar en la penumbra.
Las bacterioclorofilas y el aprovechamiento del infrarrojo
Las estrellas como nuestro Sol emiten un amplio espectro de radiación, pero la luz visible (azul y roja) es absorbida casi en su totalidad por las capas superiores de agua, las algas y la vegetación de la superficie. Si las bacterias que habitan en el fondo de un pantano dependieran de la luz roja o azul para subsistir, morirían de inanición energética de forma inmediata.
Para resolver este desafío evolutivo, estos microorganismos sintetizan bacterioclorofilas. Estos pigmentos poseen una estructura química ligeramente modificada respecto a la clorofila vegetal, lo que desplaza su espectro de absorción hacia longitudes de onda mucho más largas, situadas en la periferia de la luz roja lejana y el infrarrojo cercano (entre los 700 y los 1000 nanómetros).
La radiación infrarroja posee menos energía que la luz visible, pero tiene la propiedad de penetrar con mayor facilidad a través de las capas de lodo y los sedimentos orgánicos. De este modo, las bacterias anoxigénicas consiguen capturar un flujo de energía que es invisible y completamente inútil para las plantas superiores, explotando un nicho ecológico deshabitado en los ecosistemas profundos.
Cambios de la materia y energía durante la fotosíntesis
Centros de reacción fotosintéticos simplificados
La transferencia de la energía capturada por las bacterioclorofilas converge en estructuras proteicas denominadas centros de reacción. En el modelo oxigénico tradicional, las células coordinan dos centros de reacción distintos que operan en serie (el Fotosistema I y el Fotosistema II) para lograr la fuerza necesaria para romper el agua. La fotosíntesis anoxigénica destaca por su simplicidad estructural, ya que emplea un único fotosistema para completar todo el circuito de conversión energética.
El tipo bacteriano de tipo feofitina-quinona
Presente en las llamadas bacterias púrpuras, este centro de reacción guarda una asombrosa homología estructural con el Fotosistema II de las plantas superiores. Al recibir el impacto de los fotones, el pigmento especial se excita y expulsa un electrón de alta energía que recorre una cadena de transporte molecular basada en quinonas, impulsando el bombeo de protones a través de la membrana celular para generar energía en forma de ATP.
El tipo bacteriano de tipo hierro-azufre
Localizado en las bacterias verdes del azufre, este centro de reacción exhibe un diseño semejante al Fotosistema I de los organismos oxigénicos. En este circuito, los electrones excitados por la luz pueden tomar un camino directo para reducir coenzimas como el NADP+, proporcionando de forma inmediata el poder reductor que la célula necesita para asimilar el carbono y construir sus propias estructuras biológicas.
El club selecto de los microorganismos anoxigénicos
Este metabolismo no es universal; pertenece de forma exclusiva a un conjunto diversificado de linajes bacterianos procariontes que se dividen y clasifican en función de sus pigmentos, sus requerimientos químicos y los hábitats específicos que colonizan.
Bacterias Fotosintéticas Anoxigénicas | +--------------------------+--------------------------+ | | Bacterias Púrpuras Bacterias Verdes | | +--> Del Azufre (Chromatiaceae) +--> Del Azufre (Chlorobi) +--> No del Azufre (Rhodospirillaceae) +--> No del Azufre (Chloroflexi) Las bacterias púrpuras del azufre
Pertenecientes a la familia Chromatiaceae, estos microorganismos son habitantes fijos de zonas anóxicas (sin oxígeno) en lagos estratificados y fuentes termales ricas en compuestos de azufre. Utilizan el sulfuro de hidrógeno como donador de electrones y, al oxidarlo, acumulan gránulos de azufre elemental en el interior de su propio citoplasma. Presentan tonalidades que van desde el rosa pálido hasta el púrpura intenso debido a la combinación de sus bacterioclorofilas con pigmentos carotenoides protectores, los cuales actúan como escudos contra el daño por radiación ultravioleta.
La función de la fotosíntesis en la vida de las plantas
Las bacterias púrpuras no del azufre
Este grupo exhibe una versatilidad metabólica extraordinaria. Aunque están capacitadas para realizar la fotosíntesis anoxigénica utilizando concentraciones muy bajas de sulfuro o hidrógeno gaseoso, en ausencia de luz pueden cambiar su estrategia biológica por completo y adoptar una respiración celular heterótrofa convencional, consumiendo compuestos orgánicos del medio. Tolotan pequeñas cantidades de oxígeno, lo que les permite colonizar ambientes cambiantes donde las fronteras entre el agua óxica y la anóxica fluctúan de forma constante a lo largo del día.
Las bacterias verdes del azufre
El filo Chlorobi agrupa a microorganismos estrictamente anaerobios que perecen ante la menor traza de oxígeno libre en el ambiente. Son especialistas en la supervivencia bajo condiciones de penumbra extrema. Poseen unas estructuras colectoras de luz gigantescas denominadas clorosomas, que funcionan como parábolas satelitales ultra-sensibles, permitiéndoles realizar la fotosíntesis en zonas tan profundas del océano o de lagos negros donde la luz apenas representa una fracción insignificante de la radiación solar de la superficie. Expulsan los gránulos de azufre fuera de sus paredes celulares, alterando el aspecto del sedimento que habitan.
Las bacterias verdes no del azufre
Representadas por organismos filamentosos como Chloroflexi, estas bacterias suelen habitar en las coloridas alfombras microbianas de las fuentes geotermales de parques naturales. Prefieren utilizar compuestos orgánicos sencillos como fuente de electrones para su fotosíntesis, deslizándose de forma lenta sobre los sustratos rocosos para posicionarse en el gradiente de temperatura y radiación óptimo para su supervivencia.
Comparativa analítica de los sistemas fotosintéticos
Para fijar la frontera conceptual entre ambas modalidades de transformación energética, resulta enriquecedor contrastar sus componentes y subproductos en una estructura unificada.
La siguiente matriz detalla las diferencias fundamentales en la organización molecular de ambos metabolismos:
| Parámetro Biológico | Fotosíntesis Oxigénica | Fotosíntesis Anoxigénica |
| Donador de Electrones | Agua ({eq}\text{H}_2\text{O}{/eq}) | Sulfuro de hidrógeno, hidrógeno molecular, hierro ferroso |
| Subproducto de la Reacción | Oxígeno gaseoso ({eq}\text{O}_2{/eq}) | Azufre elemental sólido, iones sulfato o hierro férrico |
| Tipo de Clorofila | Clorofila a y b | Bacterioclorofilas (a, b, c, d, e, g) |
| Región del Espectro que Absorbe | Luz visible (400 – 700 nanómetros) | Luz roja lejana e infrarrojo (700 – 1000 nanómetros) |
| Cantidad de Fotosistemas | Dos operando en serie (PSI y PSII) | Un único fotosistema integrado |
| Organismos Representativos | Plantas, algas, cianobacterias | Bacterias púrpuras y verdes |
| Requerimiento de Oxígeno | Tolera y produce oxígeno | Estrictamente anaerobia (en la mayoría de los casos) |
La paradoja del flujo cíclico de electrones
Uno de los mecanismos más fascinantes de la fotosíntesis anoxigénica, particularmente en las bacterias púrpuras, es el uso del flujo cíclico de electrones para la producción de energía celular. Este proceso se asemeja a un circuito cerrado de generación hidroeléctrica donde el agua nunca se agota, sino que es bombeada una y otra vez a través de las mismas turbinas.
El viaje sin fin de la partícula cargada
Cuando un fotón del infrarrojo impacta en la bacterioclorofila del centro de reacción, el pigmento experimenta una alteración cuántica y expulsa un electrón de alta energía. Este electrón no se incorpora de forma inmediata a la síntesis de azúcares; en su lugar, ingresa a una cadena de transporte compuesta por transportadores lipídicos y citocromos.
A medida que el electrón desciende por esta pendiente de potencial eléctrico, su energía se aprovecha para movilizar protones hacia el exterior de la membrana celular, construyendo una diferencia de potencial idéntica a la carga de una batería. Lo verdaderamente singular es el destino final del electrón: tras agotar su energía en la cadena de transporte, un citocromo soluble recoge la partícula cargada y la devuelve exactamente al mismo centro de reacción original que la expulsó en primer lugar.
El pigmento recupera así su estado basal, quedando listo para absorber un nuevo fotón. Este movimiento circular permite a la célula generar ingentes cantidades de ATP sin necesidad de consumir de manera ininterrumpida donadores externos de electrones para la producción energética básica.
[Centro de Reacción] ---- (Electrón Excitado) ----> [Cadena de Transporte] ^ | | (Bombeo de Protones) | | +---------- (Citocromo Soluble) <-------------------+ El transporte inverso de electrones: Un esfuerzo metabólico
El flujo cíclico resuelve la demanda de energía (ATP), pero las bacterias también necesitan poder reductor (moléculas de NADH o NADPH) para fijar el dióxido de carbono y construir sus proteínas y lípidos. Aquí es donde surge un inconveniente físico: la energía de la luz infrarroja y el potencial de los electrones de las bacterias púrpuras no son suficientes para reducir directamente el NAD+.
Para solucionar este obstáculo, las bacterias ejecutan el transporte inverso de electrones. Consiste en un mecanismo donde la célula consume de forma consciente una porción del ATP que acaba de fabricar para forzar a los electrones extraídos del sulfuro de hidrógeno a viajar contracorriente, obligándolos a unirse al NAD+ para transformarlo en NADH. Este peaje energético evidencia la complejidad de subsistir en ambientes marginales empleando radiaciones de baja energía.
Una ventana al pasado geológico y la astrobiología
El estudio de la fotosíntesis anoxigénica no solo expande el conocimiento sobre la biodiversidad microbiana actual, sino que proporciona las herramientas conceptuales necesarias para descifrar la historia de nuestro planeta y buscar evidencias de vida más allá de nuestras fronteras espaciales.
El mundo antes del Gran Evento de Oxidación
Hace unos tres mil quinientos millones de años, la Tierra primitiva era un mundo radicalmente distinto al que conocemos. La atmósfera carecía de oxígeno libre y se encontraba saturada de gases como el metano, el dióxido de carbono y el nitrógeno gaseoso. En este escenario primordial, los océanos eran ricos en hierro disuelto y compuestos sulfurosos emanados del vulcanismo global.
Fueron las bacterias anoxigénicas las que dominaron los mares ancestrales durante cientos de millones de años, actuando como los principales motores de fijación de carbono del planeta. El descubrimiento de formaciones de hierro bandeado y depósitos de azufre fósil antiquísimos corrobora que estos microorganismos alteraron la química mineral de la Tierra primitiva mucho antes de que la aparición de las cianobacterias desencadenara el Gran Evento de Oxidación, una crisis climática y evolutiva que llenó la atmósfera de oxígeno y confinó a los organismos anoxigénicos a los reductos subterráneos que ocupan hoy en día.
Firmas espectrales en la búsqueda de exoplanetas
En la actualidad, los telescopios espaciales escanean las atmósferas de planetas que orbitan estrellas lejanas en busca de biomarcadores que delaten la presencia de vida extraterrestre. La presencia de oxígeno y ozono es la señal más buscada, pero los astrobiólogos advierten que un planeta habitado por organismos similares a nuestras bacterias anoxigénicas mostraría un perfil químico completamente diferente.
Si un exoplaneta orbitara alrededor de una estrella enana roja (un tipo de astro muy común en el universo que emite la mayor parte de su energía en el espectro infrarrojo), la vegetación verde sería inviable. En ese mundo, la vida prosperaría adoptando los principios de la fotosíntesis anoxigénica, cubriendo los océanos de tonalidades púrpuras o pardas y liberando gases basados en el azufre. Comprender estas rutas bioquímicas alternativas amplía los criterios de búsqueda, recordándonos que la vida puede florecer bajo cielos de cualquier color.
Resultados de aprendizaje
Al finalizar el recorrido didáctico y el análisis molecular desarrollados en este artículo de divulgación, se habrán asimilado con total claridad los siguientes conceptos:
- Definir la fotosíntesis anoxigénica como un proceso metabólico fotótrofo que utiliza compuestos distintos al agua como donadores de electrones, evitando la producción de oxígeno gaseoso.
- Explicar la adaptación cromática de las bacterioclorofilas, comprendiendo cómo su capacidad para absorber la luz infrarroja permite a estos microorganismos habitar zonas profundas desprovistas de luz visible.
- Clasificar los principales grupos de bacterias anoxigénicas, diferenciando entre bacterias púrpuras y verdes, así como sus afinidades y tolerancias respecto a los compuestos de azufre y al oxígeno.
- Describir el funcionamiento del flujo cíclico de electrones como un circuito cerrado optimizado para la síntesis de energía celular en forma de ATP.
- Reconocer el impacto histórico de estos microorganismos en la configuración geológica de la Tierra primitiva y su valor científico como modelos en el campo de la astrobiología moderna.
Referencias bibliográficas
- Brock, T. D., Madigan, M. T., Martinko, J. M., & Parker, J. (2015). Biología de los microorganismos (14.ª ed.). Pearson Educación.
- Blankenship, R. E. (2014). Molecular Mechanisms of Photosynthesis (2nd ed.). Wiley-Blackwell.
- Tortora, G. J., Funke, B. R., & Case, C. L. (2017). Introducción a la Microbiología (12.ª ed.). Editorial Médica Panamericana.
- Overmann, J., & Garcia-Pichel, F. (2013). The Phototrophic Way of Life. In The Prokaryotes (pp. 203-257). Springer, Berlin, Heidelberg.
Explora más sobre este tema
Selecciona un tema y sigue aprendiendo...
